
We discussed state dependence of noise correlations in macaque primary visual cortex [1] today. Noise correlation quantifies the covariability in spike counts between neurons (it’s called noise correlation because the signal (stimulus) drive component has been subtracted out). In a 2010 science paper [2], noise correlation was shown to be much smaller than previously reported; in the range of 0.01 compared to the usual 0.1-0.2 range and stirred up the field (see [3] for a list of values). In this paper, they argue that this difference in noise correlation magnitude is due to population level covariations during anesthesia (they used sufentanil).
They showed that a factor model with 1-dimensional latent process captures the excess noise correlations observed during anesthesia. They extended GPFA [4] to include a linear stimulus drive component, and fit the trials corresponding to the same stimulus. This model assumes that the square-root transformed spike counts in 100 ms bins 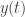


where 
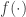

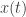

They show that the most of the extra noise correlation observed during anesthesia is explained by the 1D latent process, and the remaining noise correlation is in par with awake condition, while noise correlations in the awake data didn’t get explained away by it (figure below).

A remarkable result! (red) anesthesia, (blue) awake (solid) raw noise correlation (dashed) residual noise correlation unexplained by the 1D-latent variable
Note that this nonlinear relation between the mean rate and noise correlation is not a natural consequence of the generative model they assumed (equation above), since the noise is additive while firing rate is modulated by stimulus. That’s why they had to fit different factor loadings vector
Furthermore, they showed that local field potentials (LFP) in 0.5-2Hz range is correlated with the inferred latent variable. Hence, low-pass filtered LFP averaged over electrodes is a good proxy for L4 population correlation under anesthesia.
1D-latent process, LFP, and LNP results are all consistent with Matteo Carandini’s soloist vs chorister story except his preparations were awake rodent as far as I recall. This story also bears similarity with [5-8]. This is certainly an exciting type of model and there are many similar models being developed (e.g. [9, 10] and references therein). I hope to see more developments in these kinds of latent variable models; they have the advantage of scaling well in terms of number of parameters, but the disadvantage of being non-convex optimization. Also, I would like to see similar analysis for awake but spontaneous (non-task performing) state.
Their data and code are all available online (awesome!).
-
Ecker, A. S., Berens, P., Cotton, R. J., Subramaniyan, M., Denfield, G. H., Cadwell, C. R., Smirnakis, S. M., Bethge, M., and Tolias, A. S. (2014). State dependence of noise correlations in macaque primary visual cortex. Neuron, 82(1):235-248.
-
Ecker, A. S., Berens, P., Keliris, G. A., Bethge, M., Logothetis, N. K., and Tolias, A. S. (2010). Decorrelated neuronal firing in cortical microcircuits. Science, 327(5965):584-587.
-
Cohen, M. R. and Kohn, A. (2011). Measuring and interpreting neuronal correlations. Nat Neurosci, 14(7):811-819.
-
Yu, B. M., Cunningham, J. P., Santhanam, G., Ryu, S. I., Shenoy, K. V., and Sahani, M. (2009). Gaussian-Process factor analysis for Low-Dimensional Single-Trial analysis of neural population activity. Journal of Neurophysiology, 102(1):614-635.
-
Luczak, A., Bartho, P., and Harris, K. D. (2013). Gating of sensory input by spontaneous cortical activity. The Journal of Neuroscience, 33(4):1684-1695.
-
Goris, R. L. T., Movshon, J. A., and Simoncelli, E. P. (2014). Partitioning neuronal variability. Nat Neurosci, 17(6):858-865. [last week’s blog post]
-
Tkačik, G., Marre, O., Mora, T., Amodei, D., Berry, M. J., and Bialek, W. (2013). The simplest maximum entropy model for collective behavior in a neural network. Journal of Statistical Mechanics: Theory and Experiment, 2013(03):P03011+.
-
Okun, M., Yger, P., Marguet, S. L., Gerard-Mercier, F., Benucci, A., Katzner, S., Busse, L., Carandini, M., and Harris, K. D. (2012). Population rate dynamics and multineuron firing patterns in sensory cortex. The Journal of neuroscience : the official journal of the Society for Neuroscience, 32(48):17108-17119.
-
Buesing, L., Sahani, M., and Macke, J. H. (2012). Spectral learning of linear dynamics from generalised-linear observations with application to neural population data. NIPS 25
-
Pfau, D., Pnevmatikakis, E. A., and Paninski, L. (2013). Robust learning of low-dimensional dynamics from large neural ensembles. NIPS 26
